Crataegus
See list of 47 species in this genusCrataegus is taxonomically complex and, as treated here, the second largest genus of tracheophytes in New England. Though the genus does suffer from real phenomena, such as infrequent hybridization, apomixis, and rarely collected (i.e., poorly known) taxa, the major hurdle to learning the genus is the name proliferation by early hawthorn researchers. The following key will work best with flowering material for most species, though some taxa will require mature fruits as well for confident identification. Most measurements were performed on dried specimens, so adjustments may be needed for freshly collected material. Species with moderate to dense pubescence in the inflorescence during flowering usually maintain a few hairs along the peduncle and/or pedicels in fruit. Species with pubescent fruits also showed pubescent hypanthia during flowering. Species with one cycle of stamens have 5–10 (–11) total stamens, whereas those with two cycles of stamens have 12–20 total stamens. When assessing the character states of bracteoles within the inflorescence, those at the very base of the inflorescence should not be used as they are often different from those in the middle and upper portions of the inflorescence (they are often toothed or lobed and sometimes of herbaceous texture when the upper are not). Inflorescences/infructescences are always borne on short shoots, but not all short shoots bear flowers/fruits. Leaves from long shoots are usually larger, with more pronounced lobes, and/or broader at the base than short shoot leaves. Erosions on the inner surfaces of the pyrenes are a critical character for the identification of Crataegus; however, these are sometimes caused by insect feeding within the immature pomes. Pomes that contain living insect larvae, have insect exit holes on the exterior of the fruit, have insect feces within the erosions, or have erosions that lead to the outer surface of the pyrene should be considered suspect (i.e., find pomes that do not possess any of these features). The distribution reports for New England Crataegus are hopelessly confused by misidentifications and equivocal specimens. Except for selected species (e.g., those of regional conservation concern), no effort has been made to list all of the erroneous reports. The report of C. magniflora Sarg. from VT by Macklin (2001) is based on a collection C. coccinioides (specimen at VT!). References: Palmer (1952), Phipps and Muniyamma (1980).
The following are imperfectly known taxa:
Crataegus asperifolia Sarg. This taxon has been considered allied to C. scabrida due to the shallow and/or irregular erosions on the inner surface of some pyrenes. However, the erosions, which are very infrequent, all appear to have been caused by insects. It is known from VT.
Crataegus baccata Sarg. This hawthorn is very similar to Crataegus macracantha; however, the pomes are very small and pyrenes are plane on the inner surfaces. It is known from MA.
Crataegus bellula Sarg. This taxon has been considered to be an interserial hybrid between series Intricatae (e.g., C. intricata) and series Pruinosae (e.g., C. pruinosa). It is known from MA.
Crataegus bristolensis Sarg. This taxon is very close to C. populnea in many important morphological details. Its pyrenes show irregular erosions on the inner surfaces; however, these appear to be caused by insect damage. It is known from MA.
Crataegus cyclophylla Sarg. This taxon has been considered allied to C. scabrida due to the shallow and/or irregular erosions on the inner surface of some pyrenes. However, the erosions, which are very infrequent, all appear to be caused by insects. Leaf blade morphology suggests this is an interserial hybrid between series Rotundifoliae (e.g., C. chrysocarpa) and series Tenuifoliae (e.g., C. macrosperma). It is known from VT.
Crataegus emersoniana Sarg. This hawthorn appears to be an interserial hybrid derived from series Macracanthae (e.g., C. macracantha), as evidenced by the shallow and/or irregular erosions on the inner surface of some pyrenes. The other possible parent has not been determined with confidence but may involve series Pruinosae (e.g., C. pruinosa). It is known from MA.
Crataegus pilosa Sarg. This taxon has been considered to be an interserial hybrid between series Intricatae (e.g., C. intricata) and series Pruinosae (e.g., C. pruinosa). However, key aspects of its morphology contradict this hypothesis, including the absence of bracteoles in the inflorescence. It is known from MA.
Crataegus thayeri Sarg. This taxon is related to the C. scabrida complex but appears to be transitional between some taxa in regard to its leaf blade outline. It is known from MA.
Crataegus websteri Sarg. This hawthorn is similar to C. egglestonii in many key features and likely has a similar origin—as a hybrid taxon involving series Macracanthae (e.g., C. macracantha). It is known from NH.
-
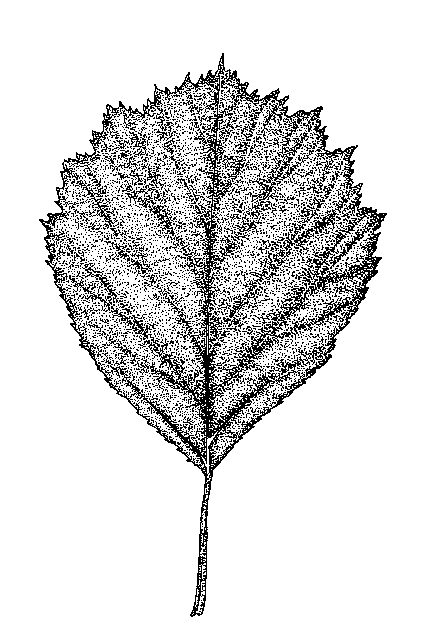
1a. Primary lateral veins of the leaf blades running to the sinuses as well as the points of the lobes [Fig. 836]
-
2a. Flowers with 3–5 styles; leaf blades rounded to subcordate at base, sharply acute at the apex [Fig. 836]; anthers non-anthocyanic; thorns (12–) 23–55 mm long
-
2b. Flowers with 1 style; leaf blades cuneate to broad-cuneate at base, rounded to acute at apex; anthers anthocyanic; thorns 5–17 mm long
-
-
1b. Primary lateral veins of the leaf blades running only to the points of the lobes or the leaves unlobed and the veins running to the teeth [Fig. 833]
-
3a. Androecium with 2 cycles of stamens [Fig. 838]
-
4a. Inflorescence sparsely to densely pubescent, retaining some hairs at least until full maturity of pome
-
5a. Flowers 20–24 mm wide; leaf blades broad-ovate to broad-triangular or suborbicular and (50–) 61–110 mm long at maturity; pomes usually prominently glaucous
-
5b. Flowers 13–20.5 mm wide; leaf blades usually of narrower shape or of shorter length or both; pomes not or only slightly glaucous
-
6a. Sepals entire or nearly so; leaf blades elliptic to narrow-elliptic-obovate to obovate, usually widest above the middle, barely (if at all) lobed on short shoots, 1.4–2.1 times as long as wide
-
6b. Sepals glandular-serrate to glandular-pinnatifid; leaf blades not combining the above-mentioned characters
-
7a. Anthers non-anthocyanic; leaf blades near apex of long shoots suborbicular or nearly so, with small, acuminate-tipped lobes
-
7b. Anthers anthocyanic; leaf blades near apex of long shoots not combining the above mentioned characters
-
8a. Pyrenes with a definite depression occupying most of each half of the inner face (pyrenes plane or with shallow and irregular erosions in the very rare C. pisifera); anthers 0.7–1 (–1.3) mm long; inflorescence relatively large, with (7–) 12–20 flowers; leaf blades mostly rhombic to obovate, those on short shoots often with obscure lobes
-
9a. Pomes 9–13 mm in diameter; pyrenes with a deep excavation on the inner surfaces; inflorescence villous (glabrous or sparsely villous in very rare forms); flowers 15–20.5 mm in diameter (in part)
-
9b. Pomes 5–9 mm in diameter; pyrenes plane to shallowly and irregularly eroded on the inner surfaces; inflorescence glabrous to sparsely villous; flowers 13–15 mm in diameter (in part)
-
-
8b. Pyrenes plane on the inner faces; anthers 1.2–1.5 mm long; inflorescence mostly of small to medium size, with 5–12 flowers; leaf blades elliptic or ovate to elliptic-ovate or orbicular, those on short shoots with obscure to evident lobes
-
10a. Leaf blades adaxially with relatively long and soft appressed hairs mostly 0.3–0.8 mm long; short shoot leaf blades usually broad-cuneate at the base, the basal margins forming an angle of mostly less than 95 degrees; petioles frequently with a narrow, but evident, wing of green tissue, especially near the apex
-
10b. Leaf blades adaxially with relatively short and stiff appressed hairs mostly 0.2–0.4 mm long; short shoot leaf blades usually rounded at the base, the basal margins forming an angle of mostly more than 95 degrees; petioles scarcely winged
-
11a. Leaf blades broad-ovate to suborbicular (occasionally some ovate on short shoots), 1–1.2 times as long as wide [Fig. 835]; pomes globose to, infrequently, short-obloid (in part)
-
11b. Leaf blades ovate or elliptic-ovate, infrequently broad-ovate, 1.1–1.6 times as long as wide; pomes obloid to obovoid
-
-
-
-
-
-
-
4b. Inflorescence glabrous
-
12a. Pyrenes with a definite depression occupying most of each half of the inner face (pyrenes plane or with shallow and irregular erosions in the very rare C. pisifera) [Fig. 834, R]; bracts of leaf opening strongly tinged with red to orange-red and conspicuous; inflorescence relatively large, with (7–) 12–20 flowers; sepals glandular-serrate to glandular-pinnatifid
-
13a. Pomes 9–13 mm in diameter; pyrenes with a deep excavation on the inner surfaces; inflorescence villous (glabrous or sparsely villous in very rare forms); flowers 15–20.5 mm in diameter (in part)
-
13b. Pomes 5–9 mm in diameter; pyrenes plane to shallowly and irregularly eroded on the inner surfaces; inflorescence glabrous to sparsely villous; flowers 13–15 mm in diameter (in part)
-
-
12b. Pyrenes plane on the inner faces or some faces with irregular and shallow erosions [Fig. 834, L]; bracts of leaf opening yellow-brown or weakly tinged with red, less conspicuous; inflorescence mostly of small to medium size, with 4–13 flowers; sepals entire to glandular-serrate
-
14a. Anthers (1.6–) 1.8–2.3 mm long; flowers 20–23 mm wide; pyrenes plane to shallowly and irregularly eroded on the inner surfaces
-
14b. Anthers 0.7–1.6 mm long; flowers (16–) 17–21.5 mm wide; pyrenes plane on the inner surfaces (shallowly and irregularly eroded on some pyrenes in C. brainerdii, but that species with 3 styles vs. (4–) 5 styles)
-
15a. Anthers 0.7–1 mm long; pyrenes plane to shallowly and irregularly eroded on the inner surfaces; leaf blades on short shoots mostly cuneate at the base, the basal margins forming an angle of mostly less than 95 degrees
-
15b. Anthers 1.1–1.6 mm long; pyrenes plane on the inner surfaces; short shoot leaf blades usually broad-cuneate to rounded, truncate, or subcordate at the base, the basal margins forming an angle of mostly more than 95 degrees
-
16a. Leaf blades adaxially with abundant strigae during flowering, these persisting at least along the main veins and/or in patches between the veins until early summer (in some taxa persisting nearly until maturation of the fruit); pomes not or only thinly glaucous; pedicels and short shoot branches not glaucous
-
16b. Leaf blades adaxially without strigae or the hairs sparse to moderately abundant during flowering and absent on mature, fully expanded blades; pomes moderately to prominently glaucous at maturity; pedicels and short shoot branches often thinly glaucous in late season
-
17a. Leaf blades glabrous adaxially from very early on (rarely with very few hairs); fruiting sepals prominently elevated on a collar 0.5–1.7 mm tall
-
17b. Leaf blades adaxially with sparse to moderately abundant strigae during flowering; fruiting sepals slightly elevated on a collar 0.3–0.8 mm tall
-
-
-
-
-
-
-
3b. Androecium with 1 cycle of stamens [Fig. 837]
-
18a. Bracteoles of inflorescence abundant at flowering, with prominently elevated marginal glands on stipes (0.1–) 0.2–1.4 mm long [Fig. 830]; petioles prominently glandular, sometimes even glandular-toothed; mature pomes yellow-green to bronze-green or dull red
-
19a. Foliage, inflorescence, and pome ± glabrous
-
20a. Leaf blades with 4 or 5 pairs of acute to subacuminate lobes, the tips of the lobes spreading or even slightly outcurved, those on long shoots with ovate to oblong-ovate blades; flowers 13–17 mm wide; pome obloid to obovoid, 8–13 mm thick
-
20b. Leaf blades with 3 or 4 pairs of obtuse to subacute lobes, the tips of the lobes ascending, those on long shoots with broad-ovate to suborbicular blades; flowers 17–21 mm wide; pome globose to subglobose (rarely short-obloid), 10–14 mm thick
-
-
19b. Foliage, inflorescence, and pome (at least when young) pubescent
-
21a. Anthers non-anthocyanic; pome subglobose to depressed-globose; leaf blades broad-cuneate to rounded, truncate, or subcordate at the base
-
21b. Anthers anthocyanic; pome obovoid to subglobose; leaf blades cuneate at the base
-
-
-
18b. Bracteoles of inflorescence absent to moderately abundant at flowering, with sessile to slightly elevated marginal glands on stipes as long as 0.2 mm; petioles glandular or not, but usually not glandular-toothed on short shoot leaves; mature pomes usually orange to red, often brightly so
-
22a. Inflorescence glabrous
-
23a. Petioles 2–12 (–15) mm long; leaf blades (1.2–) 1.5–2.5 times as long as wide, usually broadest at or above the middle of the blade, those on short shoots without lobes or with mostly short, obscure lobes
-
24a. Leaf blades 1.5–2.5 times as long as wide, those on short shoots usually unlobed and widest well above the middle, with faint veins on the adaxial surface (in part)
-
24b. Leaf blades 1.2–1.8 times as long as wide, those on short shoots often with short lobes and widest near or slightly above the middle, with somewhat evident veins on the adaxial surface
-
-
23b. Petioles 10–40 mm long; leaf blades 1–1.6 times as long as wide, usually broadest at or below the middle of the blade, those on short shoots with obscure to evident lobes
-
25a. Leaf blades adaxially with abundant strigae during flowering, the hairs persisting at least along the main veins and/or in patches between the veins until early summer (in some taxa persisting nearly until maturation of the fruit); anthers anthocyanic
-
26a. Short shoot leaves mostly cuneate at the base during flowering, the basal margins forming an angle of less than 90 degrees; pyrenes plane to shallowly and irregularly eroded on the inner surfaces (in part)
-
26b. Short shoot leaves mostly broad-cuneate to rounded or truncate (infrequently subcordate) at the base, the basal margins forming an angle of greater than 90 degrees; pyrenes plane on the inner surfaces
-
27a. Sepals entire or weakly glandular-serrate; flowers 14–16 mm wide
-
27b. Sepals glandular-serrate, often prominently so; flowers 16–20 mm wide
-
28a. Short shoot leaves elliptic to elliptic-ovate, 1.2–1.5 times as long as wide [Fig. 833] (in part)
-
28b. Short shoot leaves elliptic-ovate to broad-ovate or triangular-ovate, 1.1–1.3 times as long as wide
-
29a. Pomes obloid to obovoid, 8–10 mm in diameter (in part)
-
29b. Pomes globose to subglobose (rarely short-obloid), 9–12 mm in diameter (in part)
-
-
-
-
-
25b. Leaf blades adaxially glabrous or with sparse strigae during flowering and then soon glabrous (sometimes with moderately abundant strigae in C. dodgei); anthers anthocyanic or non-anthocyanic
-
30a. Sepals prominently glandular-serrate to glandular-pinnatifid with elongate calyx teeth, the longest teeth (0.3–) 0.5–1.5 mm long; flowers with 4 or 5 styles
-
30b. Sepals entire to glandular-serrate, the longest teeth (when present) 0.1–0.5 (–0.6) mm long; flowers with 2–4 (–5) styles
-
31a. Short shoot leaves mostly cuneate at the base during flowering, the basal margins forming an angle of less than 90 degrees [Fig. 831]; anthers non-anthocyanic
-
32a. Leaf blades on short shoots obtuse to subactue at the apex, with acute lobes, borne on glandular (rarely sparsely so) petioles; sepals weakly to somewhat prominently glandular-serrate
-
32b. Leaf blades on short-shoots rounded to obtuse at the apex, with rounded to obtuse lobes, borne on eglandular (rarely sparsely glandular) petioles [Fig. 832]; sepals entire to weakly glandular-serrate
-
-
31b. Short shoot leaves mostly broad-cuneate to roundeded or truncate (infrequently subcordate) at the base, the basal margins forming an angle of greater than 90 degrees; anthers anthocyanic (non-anthocyanic in some C. handyae, but that species with erosions on the inner surfaces of some pyrenes)
-
33a. Inner surface of some pyrenes with shallow and irregular erosions; anthers usually non-anthocyanic (in part)
-
33b. Inner surface of pyrenes plane; anthers anthocyanic
-
34a. Sepals glandular-serrate; branch thorns very fine, usually 1.6–1.7 mm wide at the midpoint; pomes obloid to obpyriform
-
34b. Sepals entire to weakly glandular-serrate; branch thorns thicker, (1.7–) 1.8–2.5 mm thick at the midpoint; pomes globose to subglobose (to obloid or obovoid in some C. populnea, but that species sparsely pubescent adaxially during flowering)
-
35a. Leaf blades adaxially with sparse to moderately abundant strigae during flowering and then soon glabrous; pomes variable in shape, sometimes elongate and tapering to the base in drying
-
35b. Leaf blades adaxially glabrous or with very few hairs during flowering; pomes globose to subglobose, rounded to truncate at the base in drying
-
36a. Leaf blades with relatively shallow sinuses, the lobes near the middle of the blade on mature leaves 4–8 mm long
-
36b. Leaf blades with relatively deep sinuses, the lobes near the middle of the blade on mature leaves 7–15 mm long
-
-
-
-
-
-
-
-
-
22b. Inflorescence sparsely to densely villous
-
37a. Pyrenes with a definite depression occupying most of each half of the inner face [Fig. 834, R]; bracts of leaf opening strongly tinged with red to orange-red and conspicuous; inflorescence relatively large, with (8–) 10–31 flowers; sepals conspicuously glandular-serrate to glandular-pinnatifid; leaf blades mostly rhombic to obovate, those on short shoots often with inconspicuous lobes
-
37b. Pyrenes plane on the inner faces or some faces with irregular and shallow erosions [Fig. 834, L]; bracts of leaf opening yellow-brown or weakly tinged with red, less conspicuous; inflorescence mostly of small to medium size, with 3–16 (–19) flowers; sepals entire or weakly to conspicuously glandular-serrate; leaf blades rhombic or elliptic to ovate, suborbicular, or elliptic-ovate (rarely oblong-obovate), those on short shoots with inconspicuous to evident lobes
-
38a. Leaf blades adaxially glabrous or with very sparse strigae during flowering and then soon glabrous
-
39a. Leaf blades oblanceolate to broad-oblanceolate-elliptic, 1.5–2.5 times as long as wide, borne on petioles 2–12 (–15) mm long, those on short shoots essentially unlobed (in part)
-
39b. Leaf blades broad-rhombic or broad-elliptic to ovate, rhombic-ovate broad-ovate, or triangular-ovate, 1–1.3 times as long as wide, borne on petioles 17–35 mm long, those on short shoots with evident lobes
-
40a. Flowers 19–22 mm wide; pomes globose; some pyrenes with shallow and irregular erosions in the inner surfaces (in part)
-
40b. Flowers 14–16 mm wide; pomes obovoid to obpyriform; pyrenes plane on the inner surfaces
-
-
-
38b. Leaf blades adaxially with abundant strigae during flowering, the hairs persisting at least along the main veins and/or in patches between the veins until early summer (in some taxa persisting nearly until maturation of the fruit)
-
41a. Short shoot leaves mostly cuneate at the base during flowering, the basal margins forming an angle of less than 95 degrees
-
42a. Flowers 19–22 mm wide; sepals entire
-
42b. Flowers 13–19 mm wide; sepals weakly to conspicuously glandular-serrate
-
43a. Pyrenes plane on the inner surfaces [Fig. 834, L]; anthers non-anthocyanic (anthocyanic in the very rare C. chrysocarpa var. praecox)
-
44a. Inflorescence sparsely pubescent; hypanthium glabrous or sparsely pubescent near the base; sepals weakly glandular-serrate
-
44b. Inflorescence moderately to densely pubescent; hypanthium pubescent ± throughout; sepals conspicuously glandular-serrate
-
45a. Leaf blades broad-rhombic to suborbicular, 36–61 mm long at maturity, 1–1.2 times as long as wide [Fig. 831]; pomes globose
-
45b. Leaf blades rhombic-elliptic to ovate, 48–78 (–87) mm long at maturity, 1.2–1.7 (–1.9) times as long as wide; pomes subglobose to short-obloid
-
-
-
43b. Some pyrenes with shallow and irregular erosions on the inner surfaces; anthers anthocyanic
-
46a. Inflorescence moderately villous; hypanthium moderately villous; young branchlets sparsely villous; flowers 18–19 mm wide; pomes obloid to obovoid, sparsely villous; leaf blades elliptic to rhombic or rhombic-ovate, usually widest near the middle, the margins with ± straight taper to the apex
-
46b. Inflorescence sparsely pubescent; hypanthium glabrous or sparsely pubescent; young branchlets glabrous; flowers 14–18 mm wide; pomes globose to subglobose (rarely to short-obloid), glabrous; leaf blades very variable in shape but usually with one or more features: somewhat oblong in outline; broadest above or below the middle; and/or the margins with concave taper to the apex
-
47a. Branch thorns long, slender, and straight or nearly so, (35–) 45–60 ×1.9–2.1 mm
-
47b. Branch thorns variable, usually shorter, more stout, and slightly curved, 22–45 (–65) × (1.9–) 2–2.9 mm
-
48a. Most leaf blades on short shoots elliptic to elliptic-obovate to oblong-obovate, with concave taper to the apex; hypanthium usually glabrous (in part)
-
48b. Most leaf blades on short shoots ovate to rhombic-ovate or broad-ovate, with convex or straight taper to the apex; hypanthium usually sparsely villous near the base
-
-
-
-
-
-
41b. Short shoot leaves mostly broad-cuneate to rounded or truncate (infrequently subcordate) at the base, the basal margins forming an angle of greater than 95 degrees
-
49a. Anthers non-anthocyanic; subherbaceous bracteoles sometimes present in the apical half of the inflorescence; stamens ca. 50% as long as the petals; leaf blades permanently pubescent abaxially on and between the primary lateral veins
-
50a. Flowers 20–26 mm wide; mature leaf blades 55–111 mm long
-
50b. Flowers 16–17.5 mm wide; mature leaf blades 36–70 mm long
-
-
49b. Anthers anthocyanic; herbaceous bracteoles rarely present in the apical half of the inflorescence; stamens ca. 65–95% as long as the petals; leaf blades abaxially pubescent on only the primary lateral veins at maturity (if at all; sometimes permanently pubescent across the surface in C. holmesiana, but that species usually with obloid to obovoid pomes vs. globose to short-obloid pomes)
-
51a. Sepals subentire to weakly glandular-serrate (i.e., sepals without marginal glands or these present and then usually sparse and mostly not elevated from the margin more than 0.2 mm), somewhat persistent or deciduous from mature pomes; flowers 15–18 mm wide
-
52a. Leaf blades broad-ovate to suborbicular (rarely some ovate or broad-triangular-ovate), 1–1.2 times as long as wide (in part)
-
52b. Leaf blades ovate to broad-elliptic or elliptic-ovate, (1.1–) 1.2–1.7 times as long as wide
-
-
51b. Sepals glandular-serrate, often conspicuously so (i.e., some of the glands elevated from the margin of the sepal on teeth 0.2–0.6 mm), somewhat to firmly persistent on the mature pomes; flowers (16–) 17–21 mm wide
-
53a. Mature leaf blades (i.e., post-flowering) on short shoots mostly elliptic to elliptic-ovate, 1.2–1.5 times as long as wide [Fig. 833] (in part)
-
53b. Mature leaf blades on short shoots ovate to suborbicular, 1–1.3 times as long as wide
-
54a. Pomes obloid to obovoid; hypanthium glabrous; lobes of the leaf blades with relatively more frequent and more prominent outcurved tips [Fig. 835] (in part)
-
54b. Pomes globose to subglobose (rarely to short-obloid); hypanthium villous (sometimes glabrous in C. coccinea); lobes of the leaf blades with relatively fewer and less prominent outcurved tips
-
55a. Leaf blades ovate to broad-ovate, usually truncate to subcordate at the base, ± plane in life; thorns (20–) 25–60 mm long (in part)
-
55b. Leaf blades broad-ovate to suborbicular, usually rounded at the base, often convex in life (i.e., with drooping sides); thorns 15–35 (–42) mm long
-
-
-
-
-
-
-
-
-
-
-
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
![]() Show photos of:
Each photo represents one species in this genus.
Show photos of:
Each photo represents one species in this genus.